

Если у вас возник любой вопрос по сайту или продукту, мы с радостью ответим вам по телефону или электронной почте.
СОСТОЯНИЕ И ПУТИ РАЗВИТИЯ АВКВАКУЛЬТУРЫ В
РОССИЙСКОЙ ФЕДЕРАЦИИ
РОССИЙСКОЙ ФЕДЕРАЦИИ
МИНИСТЕРСТВО СЕЛЬСКОГО ХОЗЯЙСТВА РОССИЙСКОЙ ФЕДЕРАЦИИ
ФЕДЕРАЛЬНОЕ АГЕНТСТВО ПО РЫБОЛОВСТВУ
ФГБОУ ВО «КАЛИНИНГРАДСКИЙ ГОСУДАРСТВЕННЫЙ ТЕХНИЧЕСКИЙ УНИВЕРСИТЕТ»
ФГБОУ ВО «САРАТОВСКИЙ ГОСУДАРСТВЕННЫЙ АГРАРНЫЙ УНИВЕРСИТЕТ им. Н.И. ВАВИЛОВА»
ФЕДЕРАЛЬНОЕ АГЕНТСТВО ПО РЫБОЛОВСТВУ
ФГБОУ ВО «КАЛИНИНГРАДСКИЙ ГОСУДАРСТВЕННЫЙ ТЕХНИЧЕСКИЙ УНИВЕРСИТЕТ»
ФГБОУ ВО «САРАТОВСКИЙ ГОСУДАРСТВЕННЫЙ АГРАРНЫЙ УНИВЕРСИТЕТ им. Н.И. ВАВИЛОВА»
V НАЦИОНАЛЬНАЯ НАУЧНО-ПРАКТИЧЕСКАЯ КОНФЕРЕНЦИЯ
Редакционная коллегия:
Васильев А.А., Кузнецов М.Ю., Руднева О.Н., Сивохина Л.А.
Васильев А.А., Кузнецов М.Ю., Руднева О.Н., Сивохина Л.А.
ПОКАТНАЯ МИГРАЦИЯ ЛИЧИНОК В Р.ВЕРХНЯЯ АНГАРА, КАК ЭЛЕМЕНТ ФУНКЦИОНАЛЬНЫХ АДАПТАЦИЙ В ВОСПРОИЗВОДСТВЕННОМ ЦИКЛЕ ОМУЛЯ СЕВЕРОБАЙКАЛЬСКОЙ ПОПУЛЯЦИИ
Материал V Национальной научно-практической конференции
"Состояние и пути развития аквакультуры в Российской Федерации"
Авторы опубликованного материала несут ответственность за некорректное заимствование, подбор и точность приведенных фактов, цитат, статистических данных и материалов не подлежащих открытой публикации. Материал размещен в авторской редакции.
"Состояние и пути развития аквакультуры в Российской Федерации"
Авторы опубликованного материала несут ответственность за некорректное заимствование, подбор и точность приведенных фактов, цитат, статистических данных и материалов не подлежащих открытой публикации. Материал размещен в авторской редакции.
М.Г. ВОРОНОВ
M.G. Voronov
БГСХА им. В.Р. Филиппова, н.с. Байкальского филиала ФГБНУ «ВНИРО»
V. Р. Filippov bssa, head of the Baikal branch OF VNIRO
M.G. Voronov
БГСХА им. В.Р. Филиппова, н.с. Байкальского филиала ФГБНУ «ВНИРО»
V. Р. Filippov bssa, head of the Baikal branch OF VNIRO
УДК: 639.3
Аннотация. Приводятся данные по покатной миграции личинок байкальского омуля с естественных нерестилищ р. Верхняя Ангара, её притоков первого порядка в 2017 году от производителей с аномально поздним сроком захода 2016 года. Установлено, что скат личинок омуля в р.Верхняя Ангара приурочен к периоду экологического оптимума. Предлагается схема промеров наиболее информативных морфологических показателей личинок омуля на первых этапах постэмбрионального развития. Определены и рассмотрены причины различий распределения, скатывающихся личинок омуля на нагул с р.Кичера и р. Верхняя Ангара в Северобайкальском соре и динамики их выхода в оз.Байкал. Показана взаимосвязь особенностей нерестовой миграции производителей омуля, закономерностям освоения ими нерестилищ в р.Верхняя Ангара и ее притоках.и покатной миграции личинок. Установлено, что механизм экологической обусловленности покатной миграции личинок байкальского омуля является элементом функциональных адаптаций в воспроизводственном цикле популяции анадромных мигрантов.
Ключевые слова: омуль, личинки, покатная миграция, морфогруппа, экологический оптимум.
Abstract. Data on the downstream migration of the Baikal omul larvae from the natural spawning grounds of the river are presented. Upper Angara, its tributaries of the first order in 2017 from producers with an abnormally late date of entry 2016. It was found that the skate of omul larvae in the Verkhnyaya Angara River is confined to the period of ecological optimum. The scheme of measurements of the most informative morphological parameters of omul larvae at the first stages of postembryonic development is proposed. The reasons for the differences in the distribution of migrating omul larvae for feeding from the Kichera River and the Upper Angara in the Severobaikalsk Sor and the dynamics of their outflow into Lake Baikal. The relationship between the features of spawning migration of omul spawners, the patterns of their development of spawning grounds in the Verkhnyaya Angara River and its tributaries, and the downstream migration of larvae are shown. It has been established that the mechanism of ecological conditioning of the downstream migration of the Baikal omul larvae is an element of functional adaptations in the reproductive cycle of the population of anadromous migrants.
Key words: omul, larvae, downstream migration, morphogroup, ecological optimum.
Введение
Учетные работы по скату личинок омуля в основных нерестовых реках Байкала в мониторинговом режиме начались в 70-х годах. В то время исследователи ставили своей задачей учета количества скатывающихся личинок, отслеживание динамики ската в зависимости от температуры воды, воздуха и уровня воды и изменение их линейно-весовых показателей. Фактически все имеющиеся публикации по скату личинок омуля констатируют ежегодные данные по количеству ежегодно скатывающихся личинок, динамики этих показателей и их взаимосвязь с температурой и уровнем воды (Краснощеков, 1958; Мишарин, 1953; 1965; Афанасьев, 1980; 1981; Сорокин, 1981; 1977; 1978;1981; Шулев, 1982; Шербаков, 1983). Базов А.В приводит наиболее полную библиографическую сводку с приведением авторских данных некоторых количественных показателей по скату и нагулу личинок омуля в разных реках Байкала ( Базов, Базова, 2016). Попытка рассмотрения функциональной роли покатной миграции личинок омуля и ее взаимосвязь с условиями нагула была предпринята Шумиловым (Шумилов, 1971; 1974) и в некоторой степени Сорокиным В.Н. (Сорокин, 1981; Сорокин, Сорокина, 1977).
Благодаря разработанной автором в 1987 году экспересс-методике разделения личинок омуля с естественных нерестилищ р.Селенги и личинок омуля, выпускаемых с Селенгинского экспериментального рыбоводного завода (СЭРЗ) (Воронов, 1993), в 1987-91 гг. было установлено, что большая часть личинок от искусственного воспроизводства элиминирует в период ската по реке. За время интенсивной работы СЭРЗ с 1981 по 1995 общий скат личинок омуля с р.Селенги упал более чем в 2 раза. К 90м годам доля омуля пелагической морфогруппы в уловах снизилась на фоне падения общего вылова омуля на Селенгинском мелководье и Малом Море. Хотя еще ранее в 50х годах исследователи аргументировано указывали на несовершенство биотехники искусственного воспроизводства байкальского омуля, а также о необходимости приведения ее в соответствие каждого этапа таковым процессам в естественных условиях (Подлесный, 1955; Черняев, 1968; 1982). Основной целью данной работы является выяснение функции покатной миграции личинок омуля в период воспроизводства и ее экологической обусловленности.
Материал и методика
Основой данной работы послужили материалы, собранные автором при изучении покатной миграции и учете численности личинок байкальского омуля с верхних нерестилищ в реке Верхняя Ангара, результаты дважды проведенной траловой съемки Северобайкальского сора весной 2017 года, а также исследования по скату личинок омуля с разных участков нерестилищ р.Селенги и ее притоков с 1983-2007гг., по динамике распределения (выхода) личинок омуля на нагул в дельте Селенги и Селенгинском мелководье, Посольском соре, в устьевых пространствах рр.Кика, Турка, Безымянка, Большой и Малый Чивыркуй в 1986-2002гг. . Основной створ учета личинок омуля был в р.Верхняя Ангара выше устья реки Окушанда, дополнительно контрольные пробы брались в устье реки Котера, в р.Верхняя Ангара выше устья р,Котера и выше около 1.5 км основного створа учета. Также в 2017 году проведен учет личинок омуля и в левом притоке р.Верхняя Ангара - р.Окушанда. Учет личинок омуля велся в соответствии с общепризнанной для омуля методикой повременным способом (Шумилов,1974; Сорокин,1977; Афанасьев, 1981; Воронов, 1993). Сущность метода заключается в регулярном взятии проб на трёх постоянных станциях по трем горизонтам, где были выставлены буи на «мертвяках». Таким образом, при каждом взятии проб охватывался весь профиль реки. Для выявления суточной динамики ската личинок регулярно, раз в три дня, пробы брали круглосуточно через 4 часа. Экспозиция постановки сеток Раса-Мишарина зависела от интенсивности ската и составляла от 1 до 10 минут. Конструкция этих сеток была несколько изменена – цилиндрическая часть из бязи была заменена обратным конусом из легкого брезента, что существенно увеличило фильтрующую часть ловушки. Такая конструкция позволяет сохранять коэффициент фильтрации (коэффициент уловистости) равным единице, даже при увеличении скорости реки и мутности речного потока (Воронов, 1993). Каждые трое суток отбиралась проба личинок, которых фиксировали в смеси 2-х % формалина и 70% спирта на физрастворе в соотношении 1:1. Такая фиксирующая смесь не приводит к изменению морфометрических параметров личинок. Одновременно на свежем материале по отработанной ранее схеме проводились промеры 9 параметров (Воронов, 1998) (Рис.1).
Ключевые слова: омуль, личинки, покатная миграция, морфогруппа, экологический оптимум.
Abstract. Data on the downstream migration of the Baikal omul larvae from the natural spawning grounds of the river are presented. Upper Angara, its tributaries of the first order in 2017 from producers with an abnormally late date of entry 2016. It was found that the skate of omul larvae in the Verkhnyaya Angara River is confined to the period of ecological optimum. The scheme of measurements of the most informative morphological parameters of omul larvae at the first stages of postembryonic development is proposed. The reasons for the differences in the distribution of migrating omul larvae for feeding from the Kichera River and the Upper Angara in the Severobaikalsk Sor and the dynamics of their outflow into Lake Baikal. The relationship between the features of spawning migration of omul spawners, the patterns of their development of spawning grounds in the Verkhnyaya Angara River and its tributaries, and the downstream migration of larvae are shown. It has been established that the mechanism of ecological conditioning of the downstream migration of the Baikal omul larvae is an element of functional adaptations in the reproductive cycle of the population of anadromous migrants.
Key words: omul, larvae, downstream migration, morphogroup, ecological optimum.
Введение
Учетные работы по скату личинок омуля в основных нерестовых реках Байкала в мониторинговом режиме начались в 70-х годах. В то время исследователи ставили своей задачей учета количества скатывающихся личинок, отслеживание динамики ската в зависимости от температуры воды, воздуха и уровня воды и изменение их линейно-весовых показателей. Фактически все имеющиеся публикации по скату личинок омуля констатируют ежегодные данные по количеству ежегодно скатывающихся личинок, динамики этих показателей и их взаимосвязь с температурой и уровнем воды (Краснощеков, 1958; Мишарин, 1953; 1965; Афанасьев, 1980; 1981; Сорокин, 1981; 1977; 1978;1981; Шулев, 1982; Шербаков, 1983). Базов А.В приводит наиболее полную библиографическую сводку с приведением авторских данных некоторых количественных показателей по скату и нагулу личинок омуля в разных реках Байкала ( Базов, Базова, 2016). Попытка рассмотрения функциональной роли покатной миграции личинок омуля и ее взаимосвязь с условиями нагула была предпринята Шумиловым (Шумилов, 1971; 1974) и в некоторой степени Сорокиным В.Н. (Сорокин, 1981; Сорокин, Сорокина, 1977).
Благодаря разработанной автором в 1987 году экспересс-методике разделения личинок омуля с естественных нерестилищ р.Селенги и личинок омуля, выпускаемых с Селенгинского экспериментального рыбоводного завода (СЭРЗ) (Воронов, 1993), в 1987-91 гг. было установлено, что большая часть личинок от искусственного воспроизводства элиминирует в период ската по реке. За время интенсивной работы СЭРЗ с 1981 по 1995 общий скат личинок омуля с р.Селенги упал более чем в 2 раза. К 90м годам доля омуля пелагической морфогруппы в уловах снизилась на фоне падения общего вылова омуля на Селенгинском мелководье и Малом Море. Хотя еще ранее в 50х годах исследователи аргументировано указывали на несовершенство биотехники искусственного воспроизводства байкальского омуля, а также о необходимости приведения ее в соответствие каждого этапа таковым процессам в естественных условиях (Подлесный, 1955; Черняев, 1968; 1982). Основной целью данной работы является выяснение функции покатной миграции личинок омуля в период воспроизводства и ее экологической обусловленности.
Материал и методика
Основой данной работы послужили материалы, собранные автором при изучении покатной миграции и учете численности личинок байкальского омуля с верхних нерестилищ в реке Верхняя Ангара, результаты дважды проведенной траловой съемки Северобайкальского сора весной 2017 года, а также исследования по скату личинок омуля с разных участков нерестилищ р.Селенги и ее притоков с 1983-2007гг., по динамике распределения (выхода) личинок омуля на нагул в дельте Селенги и Селенгинском мелководье, Посольском соре, в устьевых пространствах рр.Кика, Турка, Безымянка, Большой и Малый Чивыркуй в 1986-2002гг. . Основной створ учета личинок омуля был в р.Верхняя Ангара выше устья реки Окушанда, дополнительно контрольные пробы брались в устье реки Котера, в р.Верхняя Ангара выше устья р,Котера и выше около 1.5 км основного створа учета. Также в 2017 году проведен учет личинок омуля и в левом притоке р.Верхняя Ангара - р.Окушанда. Учет личинок омуля велся в соответствии с общепризнанной для омуля методикой повременным способом (Шумилов,1974; Сорокин,1977; Афанасьев, 1981; Воронов, 1993). Сущность метода заключается в регулярном взятии проб на трёх постоянных станциях по трем горизонтам, где были выставлены буи на «мертвяках». Таким образом, при каждом взятии проб охватывался весь профиль реки. Для выявления суточной динамики ската личинок регулярно, раз в три дня, пробы брали круглосуточно через 4 часа. Экспозиция постановки сеток Раса-Мишарина зависела от интенсивности ската и составляла от 1 до 10 минут. Конструкция этих сеток была несколько изменена – цилиндрическая часть из бязи была заменена обратным конусом из легкого брезента, что существенно увеличило фильтрующую часть ловушки. Такая конструкция позволяет сохранять коэффициент фильтрации (коэффициент уловистости) равным единице, даже при увеличении скорости реки и мутности речного потока (Воронов, 1993). Каждые трое суток отбиралась проба личинок, которых фиксировали в смеси 2-х % формалина и 70% спирта на физрастворе в соотношении 1:1. Такая фиксирующая смесь не приводит к изменению морфометрических параметров личинок. Одновременно на свежем материале по отработанной ранее схеме проводились промеры 9 параметров (Воронов, 1998) (Рис.1).
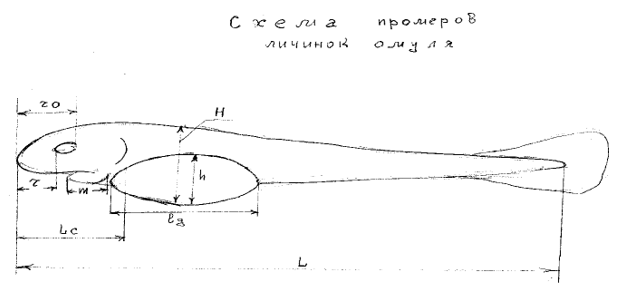
Рис.1. Схема промеров личинок омуля. Длина (L) и высота (H) тела, длина рыла (r), длина до заднего края глаза (ro), длина головы (Lc), длина нижней челюсти (m), длина (lg) и высота (h) желточного мешка, вес (Q) личинки
Результаты исследований
Скат личинок байкальского омуля в разных реках Байкала происходит в разное время и при разных условиях. В таких реках как Селенга, ее притоке Чикой, р.Большая Речка вылупление и скат личинок происходит на стадии предличинки в период вскрытия реки, т.е. весеннего ледохода. Имеющиеся данные и исследования автора свидетельствуют, что на реках, где ледоход не выражен Верхняя Ангара, Кичера, Баргузин, Кика, реки Чивыркуйского залива и ряду других рек вылупление и скат личинок с их нерестилищ происходит после вскрытия реки в период первого весеннего паводка.
Данные по скату личинок омуля с верхних нерестилищ р.Верхняя Ангара приводятся впервые. Единичный скат личинок омуля весной 2017 года начался 3 мая при температуре воды 1.5ºС при подъеме уровня воды на 23 см и приостановился к 9 мая при синхронном падении уровня воды на 17 см на фоне общего похолодания (Рис.4). Устойчиво скат личинок возобновился 11 мая, при пике интенсивности ската 19 мая. Причем 85.6% от учтенных личинок скатилось за 4 суток с 18 по 21 мая (Рис.2 и 3), при общем скате 132.28 млн.шт.
Результаты исследований
Скат личинок байкальского омуля в разных реках Байкала происходит в разное время и при разных условиях. В таких реках как Селенга, ее притоке Чикой, р.Большая Речка вылупление и скат личинок происходит на стадии предличинки в период вскрытия реки, т.е. весеннего ледохода. Имеющиеся данные и исследования автора свидетельствуют, что на реках, где ледоход не выражен Верхняя Ангара, Кичера, Баргузин, Кика, реки Чивыркуйского залива и ряду других рек вылупление и скат личинок с их нерестилищ происходит после вскрытия реки в период первого весеннего паводка.
Данные по скату личинок омуля с верхних нерестилищ р.Верхняя Ангара приводятся впервые. Единичный скат личинок омуля весной 2017 года начался 3 мая при температуре воды 1.5ºС при подъеме уровня воды на 23 см и приостановился к 9 мая при синхронном падении уровня воды на 17 см на фоне общего похолодания (Рис.4). Устойчиво скат личинок возобновился 11 мая, при пике интенсивности ската 19 мая. Причем 85.6% от учтенных личинок скатилось за 4 суток с 18 по 21 мая (Рис.2 и 3), при общем скате 132.28 млн.шт.
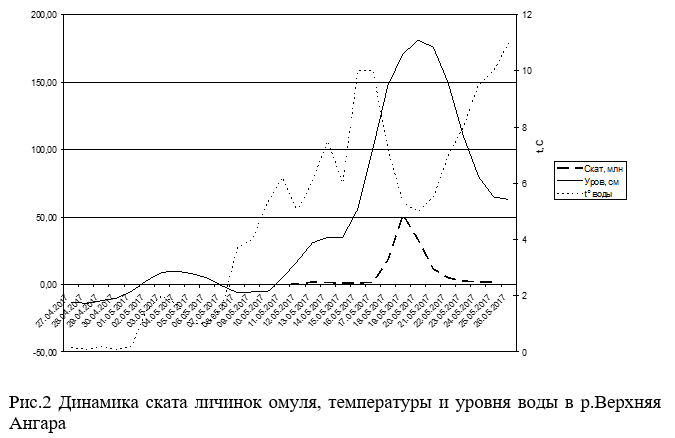
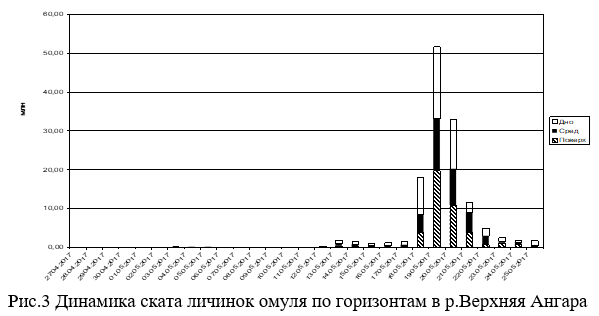
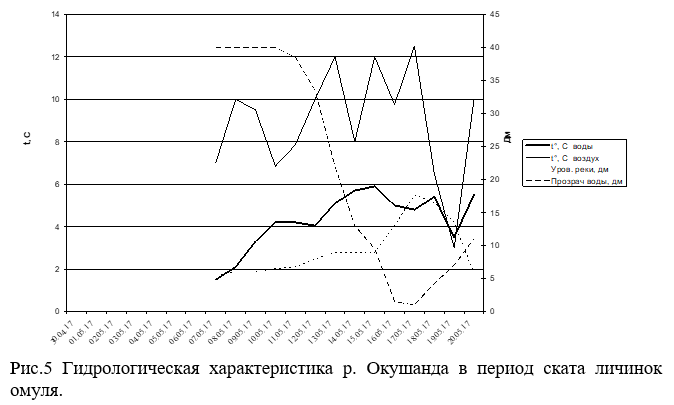
Единичный скат личинок в р.Окушанда начался 13 мая при максимуме 15 мая в общей сложности скатилось 2.69 млн.шт. Зафиксировано аномальное явление массового сноса мертвой икры. До подъема уровня воды, где была возможность пройти на лодке, а это около 1.5 км от устья, на дне были обнаружены наполовину разложившие трупы производителей омуля. Были вскрыты около полутора десятков уцелевших трупов, все они были отнерестившимися. Из этого следует, что гибель их произошла после нереста из-за перемерзания русла реки. Ток воды в период обследования нерестилищ р.Окушанда 27-30 апреля практически отсутствовал. Основной причиной промерзания русла реки можно считать устройство стихийной переправы для вывоза леса с делянок, отведенных по берегам этой нерестовой реки. Мертвая икра стала попадаться в пробах также 13 мая при пике сноса 17 мая, который состоялся за сутки до пика подъема уровня воды. В массе мертвую икру несло в придонном (более 85%) горизонте. Всего учтено 150.89 млн. мертвых икринок.
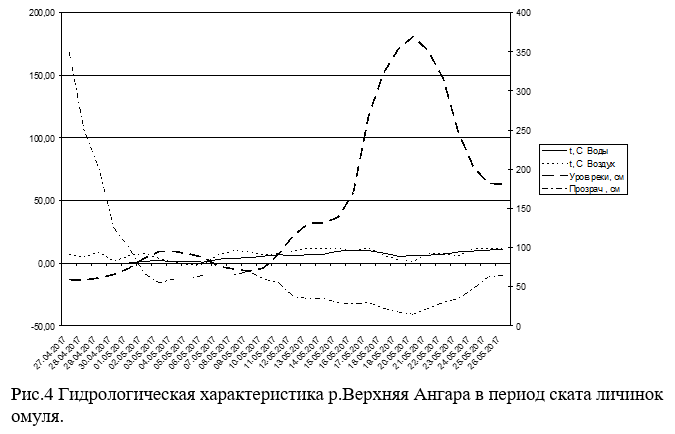
Данные гидрологических показателей, как реки Верхней Ангары, так и реки Окушанда (Рис.4 и 5) свидетельствуют, что основной скат личинок происходил во время максимального подъема уровня реки (до 2м), при резком падении прозрачности (с 3.5м до 0.25-0.3м) и температуры воды (с 10°С до 3.5-5°С).
Основной скат личинок омуля 2017 г. в р.Кичера в створе ж/д моста прошел в течение одних суток 16-17 мая (по устному сообщению н.с. Кожемякина К.М Байкальского филиала ФГБНУ «Госрыбцентр» и начальника Забайкальского отдела Байкальского филиала ФГБУ «Главрыбвод» Сахарова С.В.). С 30 по 31 мая была проведена траловая съемка Северобайкальского сора по сетке станций на пяти разрезах (в общей сложности 24 траления) и трех тралений от устья р. Кичера в оз.Байкал к пирсу Нижнеангарского рыбзавода. В результате этих исследований было установлено, что после ската особенности распространения личинок омуля на нагул с р.Кичера и Верхней Ангары различны.
Данные гидрологических показателей, как реки Верхней Ангары, так и реки Окушанда (Рис.4 и 5) свидетельствуют, что основной скат личинок происходил во время максимального подъема уровня реки (до 2м), при резком падении прозрачности (с 3.5м до 0.25-0.3м) и температуры воды (с 10°С до 3.5-5°С).
Основной скат личинок омуля 2017 г. в р.Кичера в створе ж/д моста прошел в течение одних суток 16-17 мая (по устному сообщению н.с. Кожемякина К.М Байкальского филиала ФГБНУ «Госрыбцентр» и начальника Забайкальского отдела Байкальского филиала ФГБУ «Главрыбвод» Сахарова С.В.). С 30 по 31 мая была проведена траловая съемка Северобайкальского сора по сетке станций на пяти разрезах (в общей сложности 24 траления) и трех тралений от устья р. Кичера в оз.Байкал к пирсу Нижнеангарского рыбзавода. В результате этих исследований было установлено, что после ската особенности распространения личинок омуля на нагул с р.Кичера и Верхней Ангары различны.
Основная часть личинок омуля Кичерской популяции скатывается с водой первого весеннего паводка р.Кичера в Северо-Западную часть сора и частично выходит в оз.Байкал вдоль северо-западного берега. Температура воды в этой прибрежной полосе была от 10.8 до 12.3°С, в то время как в соровой части - от 9.3 до 10.4°С. В это время основная часть озера Байкал от устья р.Кичера до бухты Дагары и вдоль Восточного берега была покрыта льдом. Свободной ото льда была полоса вдоль западного берега. Личинки омуля, которые выходили в оз.Байкал были уже более развиты, у них почти полностью рассосался желточный мешок, при показателях длины тела 16.9-17.4 мм и весе 21.4-25.5 мг . Личинки омуля, скатившиеся с Верхней Ангары, в Северобайкальском соре в это время не обнаружены. Вторая основная волна весенних вод Верхней Ангары в 2017 году в створе учета личинок прокатывалась с 4 по 10 июня, причем повторный подъем воды был на 1 метр выше первого, подъем уровня воды в Северобайкальском соре происходил с 10 по 14 июня. Повторно проведенная траловая съемка показала, что личинки омуля с р.Верхняя Ангара в прибрежно-соровую систему начали поступать именно с этой волной весенних вод, т.е. с 10 июня по 14 июня. Личинки к этому времени уже достаточно развиты и организованы в стайки, по основным прорвам с током теплых вод стали выходить в оз.Байкал. К этому времени озеро полностью освободилось ото льда. Необходимо отметить, что ближе к устьям рек как при первой траловой съемке в северо-западной части сора, так и при второй в северо-восточной части сора единично попадалась молодь омуля на стадии предличинки.
Обсуждение результатов
Функция покатной миграции личинок - обеспечение оптимальных условий их нагула (Соин, 1980), уход от потенциальных хищников, что, в конечном счете, гарантирует максимальную выживаемость. Поэтому характер и протяженность покатной миграции обеспечивает благоприятные места нагула, оптимальную плотность распределения, а это обеспеченность кормом, уход от хищника, сведение к минимуму конкуренции. В свою очередь нерестовая миграция определяет протяженность и длительность покатной миграции будущего потомства. Таким образом, потомство от производителей первого периода захода на первых этапах постэмбрионального развития оказываются в наиболее благоприятных условиях нагула, по сравнению с потомством от производителей второго и тем более третьего периодов.
Так аномально поздний заход 2016 года производителей омуля в реки Верхнюю Ангару и Кичеру и наверняка с низкими миграционными возможностями, что не позволило в полной мере реализовать требуемые дальности хода для достижения наиболее благоприятных нерестилищ.
Если рассматривать покатную миграцию личинок омуля с естественных нерестилищ Верхней Ангары за прошлый период, когда количество скатывающихся личинок находилось на уровне 1 млрд. шт. и более, то максимальные значения на кривой динамики их ската были более продолжительны во времени и зачастую имели несколько пиков. То есть интенсивность ската личинок носила как бы импульсивный характер (Шумилов, 1974; Сорокин, и др., 1981; Войтов А.А., 1981; Шербаков, 1983). Как показали наблюдения 2017 года, пусть и незначительный, но скат личинок омуля с р.Котера прошел немого раньше, т.е. до основного ската личинок с нерестилищ р.В.Ангара (19-21 мая), р.Окушанда -15 мая. Исходя из анализа данных, ранее проводимых работ следует, что в р.Верхняя Ангара вылупление и скат личинок омуля происходит при первом весеннем подъеме уровня воды, которому предшествует прогрев воздуха. Проведенные нами исследования показали, что этот комплекс факторов дифференцирован для каждого притока реки обусловленных временем наступления этих условий в распадке, где протекает этот приток. В многолетнем аспекте в бассейне реки Верхняя Ангара скат личинок происходит со второй декады мая по первую декаду июня при колебаниях на 10-15 суток. Если весна дружная и подъем уровня воды в разных притоках совпадает, в нижнем течении р.Верхняя Ангара наблюдается мощный паводок и вода выходит из берегов, что отрицательно сказывается на выживаемости скатывающихся личинках омуля. Но, как правило, паводок в разных притоках наступает поочередно от устья к истоку, т.е. с Юга на Север. Поэтому и основной скат личинок омуля в каждом притоке не совпадает, что и отражается на кривой динамики интенсивности ската.
Анализ имеющих данных и наши наблюдения показывают, что численность и динамика скатывающихся личинок омуля соответствуют численности нерестового стада производителей и особенностям их при освоении нерестилищ.
С конца 90х годов в последние два десятилетия численность скатывающихся личинок омуля в Верхняя Ангара находится на уровне очень низкой и весьма стабильной величине порядка 10 % от потенциального фонда икры заходящих производителей. Столь стабильно низкая эффективность естественного воспроизводства при разном уровне водности реки кажется сомнительной. Для байкальского омуля такая эффективность воспроизводства находится не ниже критических величин (Воронов, 2003), и она не может обеспечить пополнения, гарантирующего его воспроизводительную способность. Установлено, что для каждого вида существует зависимость между индивидуальной абсолютной плодовитостью и выживаемостью потомства. Если рассматривать фазу покатной миграции личинок в воспроизводственном цикле байкальского омуля, то для обеспечения промыслового изъятия в современных условиях на уровне 20-25 тыс. ц, то среднемноголетняя величина ската личинок с р.Селенга должна быть на уровне 1.5 млрд. шт. (Воронов, 1993), для р.Верхняя Ангара 2.2-2.8 млрд. шт. (Шумилов, 1974). Хотя для байкальского омуля и установлена зависимость (Калягин, 1996; Воронов, 1993), как и для большинства видов рыб (Никольский, 1974) - выживаемости поколения от его исходной численности. Для байкальского омуля это количества скатывающихся личинок и чем ниже ее величина, тем выше выживаемость от данного поколения, но эти величины предельны. Для омуля пелагической морфогруппы минимальная критическая величина скатывающихся личинок находятся на уровне порядка 60-70% от оптимальной и не более 3-4 лет подряд, когда сохраняется отработанный режим промысла и оптимальный уровень воспроизводства (Воронов, 1993). Ярким примером этому может служить факт срыва воспроизводства омуля на р.Селенге в 1981-89 гг., когда эффективность реализации репродукционного потенциала была снижена в 2.2 раза от оптимальной величины из-за применения электро-рыбо-заградительного устройства. В натуральном выражении средняя величина скатывающихся личинок вместе с личинками выпускаемыми с Селенгинского рыбоводного завода упала в среднем до 0.72 млрд. шт, которые обеспечили заход в среднем 1.12 млн. производителей, а от этих производителей средний скат составил 0.64 млрд. личинок, из которых средний заход в р.Селенгу составил 1.02 млн. экз. Это отрицательно сказалось и на величине промыслового изъятия пелагической морфогруппы особенно на Селенгинском мелководье – его доля в уловах омуля упала с 70-80% (в 80х годах) до 40-55% (в 90х годах) обшей добычи омуля в этом промрайоне. В этот период по данным официальной статистике произошло и снижение общего вылова омуля по Селенгинскому промрайону с 800 до 300 тонн.
Поэтому резкое снижение численности скатившихся личинок омуля с р.Верхняя Ангара в 2017 году 135 млн.шт., что почти в 10 раз ниже оптимальной величины, необходимо рассматривать как катастрофическое состояние уровня воспроизводства, которое при всех суммарных компенсаторных факторах (Калягин, 1996) не в состоянии обеспечить нормальное функционирование популяции и тем более промысел.
Анализ всех природных факторов в период ската личинок омуля в р.Верхняя Ангара в 2017 году позволяет характеризовать их, как оптимальные. Вылупление и скат личинок прошли в средние сроки для этой реки при подъеме уровня воды не превышающий среднемноголетний. Поэтому выхода воды из берегов не произошло, а, следовательно, и потери скатывающихся личинок на разливах. Оптимальные сроки выхода личинок на места нагула при небольшой их численности обеспечили темп их роста несколько выше средних значений, а это позволяет надеяться на более высокую величину выживаемости омуля от поколения этого года.
По данным Войтова А.А. заход производителей омуля реки Верхняя Ангара до 2011 года был на уровне 2.3 млн.экз., потенциальный фонд икры 8.7 млрд. икринок, скат личинок около 1 млрд.шт., с 2012 г. такой же численности заходящих производителей наблюдается резкое снижение количества скатывающихся личинок почти в 2.5 раза от потенциального фонда икры и в 1,7 раза в количественном выражении. Резкое снижение количества скатывающихся личинок омуля с 2012 года произошло и на р.Селенге (Базов, 2016), хотя до последних двух лет 2016 и 2017 годах официальные данные по количеству заходящих производителей, при некотором снижении, оставались в пределах ошибки учета. Для выяснения причин столь значительного несоответствия численности заходящих производителей – количеству скатывающихся личинок и величины промыслового изъятия требуют специального анализа.
При изучении покатной миграции личинок омуля в р.В.Ангара (Шумилов, 1974) была установлена зависимость размерно-весовых показателей личинок в течение их покатной миграции – вначале скатываются самые мелкие, затем крупнее и в конце покатной миграции самые крупные, в то время как в р.Кичера в период всего ската различий в скатывающихся личинках нет. По мнению автора увеличение размера личинок происходит вследствие их развития за период ската с верхних нерестилищ, а отсутствие такового у личинок в р.Кичеры из-за близости пункта учета от нерестилищ. При изучении покатной миграции личинок омуля в р.Селенге установлено, на постоянном створе 35 км от устья вначале покатной миграции скатываются наиболее крупные личинки затем закономерно уменьшаясь как в размерном, так и весовом составе (Афанасьев 1980). В то время как, при исследовании этих показателей на удаленных друг от друга участках реки (155 км, 35 км, 2 км от устья) при относительном сохранении длины, вес личинок достоверно уменьшался (Афанасьев, 1980; Сорокин и др., 1981). Уменьшение весовых показателей по мере ската личинок авторы объясняют затратой запаса энергии при эндогенном питании.
Размеры икринок на стадии овуляции у омуля разных морфогрупп достоверно отличаются. Так наиболее крупная икра у омуля прибрежной морфогруппы, а наиболее мелкая у омуля придонно-глубоководной, среднее положение занимает икра у омуля пелагической морфогруппы. Из более крупных икринок вылупляются и более крупные предличинки. Расовая структура нерестового стада омуля каждой реки не однородна и представлена как минимум двумя морфогруппами. Исследованиями автора, проводимые по изучению нерестовых миграций омуля в реках Байкала в частности установлено, что на р.Селенге и Верхней Ангаре вначале нерестовой миграции идут наиболее крупные и сильные производители омуля пелагической морфогруппы. Это либо повторно нерестующие, либо наиболее быстрорастущие особи.
Наши исследования покатной миграции личинок также подтверждают ранее выявленные закономерности по изменению размерных показателей личинок омуля, как в реке Селенга, так и в р.Верхняя Ангара. Только вот причина этих закономерностей, что в р.Верхняя Ангара вначале скатываются более мелкие личинки, а затем более крупные, по нашему данным, обусловлены скатом личинок омуля пелагической морфогруппы, так как они находятся на более удаленных верхних нерестилищах, а затем личинки прибрежной морфогруппы. Вскрытие реки Селенга и первый весенний паводок в р Верхняя Ангара начинается с верховья каждого притока, поэтому вылупление и скат личинок начинается с наиболее верхних участков нерестилищ. В р.Верхняя Ангара вначале покатной миграции скатываются личинки омуля пелагической морфогруппы, которые более мелкие, а затем личинки омуля прибрежной морфогруппы, которые крупнее пелагических. А в р.Селенга вначале скатываются более крупные личинки от элитной части производителей пелагической морфргуппы, а затем с ниже расположенных нерестилищ от остальной части производителей, т.е. по мере вскрытия реки, а также ската личинок от придонно-глубоководной морфогруппы, которые мельче пелагических.
Из вышесказанного следует, что покатная миграция личинок омуля в каждой реке это экологически обусловленный процесс на этапе воспроизводства, как адаптивный фактор, направленный на обеспечение выживаемости с учетом особенностей воспроизводства в конкретных условиях каждой реки.
То, что сроки выклупления и характер ската личинок омуля экологически выдержаны и обусловлены, объясняет задержку скатившихся личинок омуля с нерестилищ Верхней Ангары в водоемах верхней части Северобайкальского сора. До прихода второй основной волны весеннего паводка большая Северо-Восточная часть сора была слишком мелководной (менее 50 см) с выраженным речным режимом основных проток выхода в открытый Байкал, который к тому же еще был покрыт льдом. В этот период условия для нагула личинок омуля неблагоприятны. С приходом второй волны паводка с 10 по 14 июня 2017 года условия в этой части сора стали благоприятными – поднялся уровень воды, скорость течения выноса воды из сора в озеро по основным прорвам снизилась, Байкал освободился ото льда. Непосредственно около косы и вдоль Восточного берега Байкала тоже образовалась зона теплой речной воды.
Для установления более четких и обусловленных закономерностей динамики выхода личинок из прибрежно-соровой системы в открытую часть Северного Байкала, проследить их дальнейшую миграцию и условия нагула требуются специальные исследования. Наблюдения, проведенные на фиксированных разрезах в прибрежной зоне Северной оконечности Байкала (Шумилов, 1974; Сорокин, 1977) свидетельствуют об увеличении разброса линейно-весовых показателей нагульных личинок омуля от июня к июлю. Эти факты указывают на достаточно сложный механизм приспособлений личинок омуля к разным условиям нагула в этой экосистеме. С одной стороны есть основания предполагать и наши исследования это подтверждают, что вылупление личинок на нижних участках нерестилищ происходит и в июне уже после основного ската личинок. Кроме этого личинки, а затем и молодь омуля кичерской популяции далеких миграций не совершает и продолжает нагул на северном Байкале (Смирнов, 2009), в то время как личинки и молодь ангарской популяции на нагул уходит на Юг Байкала, совершая миграции в прибрежной зоне вокруг озера. Это подтверждается тем, что уже весной в Малом Море нагуливаются годовики омуля пелагической и прибрежной морфогрупп.
Выводы
1. Основной скат личинок омуля с нерестилищ бассейна реки Верхняя Ангара в 2017 году прошел в период первого весеннего паводка с 18 по 21 мая при подъеме уровня на 194см, падении прозрачности с 3,5 м до 0.25 м, понижении температуры воды с 10.3ºС до 3.5-6ºС эти условия следует считать периодом экологического оптимума.
2. Аномально поздний заход производителей омуля 2016 года при костной нормативно-правовой базе способствовали официальному вылову производителей идущих на нерест, в результате реализованный репродукционный потенциал зашедших производителей составил 25.3%.
3. При позднем заходе производителей омуля в р.Верхняя Ангара, р.Окушанда играет существенную роль в воспроизводстве омуля, однако условия инкубации икры в 2016-17 году были нарушены и основная часть икры погибла, скат личинок составил 2.65 млн. личинок, а снос учтенных мертвых икринок150.89 млн. шт.
4. Личинки омуля из р.Кичера сразу скатываются в Северо-Западную часть Северобайкальского сора, а затем с потоком теплой соровой воды распространяется вдоль СЗ побережья оз.Байкал, освобождающуюся ото льда. Покатные личинки омуля с р.Верхней Ангары задерживаются в верховьях СВ части сора и позже при втором более мощном весеннем паводке (в 2017 году 10-14 июня), уже в более развитом состоянии выходят в СВ часть сора и с потоком теплых соровых вод выходят в прибрежную часть Северного Байкала, к этому времени освободившуюся ото льда.
5. Сроки вылупления и покатную миграцию личинок омуля экологически выдержанных и обусловленных следует рассматривать как один из элементов функциональных адаптаций в воспроизводственном цикле байкальского омуля.
6. Для установления более четких закономерностей динамики выхода и нагула личинок из прибрежно-соровой системы в открытой части Северного Байкала и дальнейших их миграций требуются специальные исследования.
Список литературы:
1. Афанасьев Г.А. Экология и воспроизводство байкальского омуля в реке Селенге: Автореф. дис. канд. биол. наук. - Иркутск, 1980. - 23 с.
2. Афанасьев Г.А. Экология нерестового стада омуля р.Селенги // Экология, болезни и разведение байкальского омуля. - Новосибирск, 1981.- С. 5-34.
3. Базов А.В., Базова Н.В. Селенгинская популяция байкальского омуля: прошлое, настоящее, будущее. – Улан-Удэ6 Изд-во БНЦ СО РАН, 2016, 352 с.
4. Войтов А.А. Биологическая характеристика и воспроизводство омуля в р.Верхняя Ангара. // Экология, болезни и разведение байкальского омуля. - Новосибирск, 1981.- С. 75-82.
5. Воронов М.Г. Эколого-биологические основы повышения эффективности воспроизводства омуля в р.Селенге в современных условиях.// Автореферат диссертации на соискание ученой степени кандидата биологических наук. - Санкт-Петербург, 1993. - 18с.
6. Воронов М.Г. Разнообразие байкальского омуля, структурная организация воспроизводящей части популяции и проблемы их сохранения.// Первый конгресс ихтиологов России: Тез. Докл. сентябрь г.Астрахань. – М., ВНИРО, 1997, С.
7. Воронов М.Г. К вопросу о перспективах искусственного воспроизводства байкальского омуля. Тез.докл. г.Улан-Удэ, 2003, С
8. Гербильский Н.Л. Биологическое значение и функциональная детерминация миграционного поведения анадромных рыб // Биологическое значение и функциональный детерминизм миграционного поведения животных. - М.; Л., 1965. - С. 18-33.
9. Дормидонтов А.С. Нерест и вопросы ленской ряпушки // Вопросы рыбного хозяйства Восточной Сибири. - Иркутск, 1969. - С. 9-19.
10. Дормидонтов А.С. Нерестовые миграции как адаптации, обеспечивающие расселение сигов по местам нагула // Зоологические исследования Сибири и Дальнего Востока: V Всесоюз. симпоз. Биологич.проблемы Севера, 1972, Магадан. - Владивосток, 1974. - С. 164 -168.
11. Краснощеков С.И. О биологии личинок байкальского омуля. //Тр./ Научно-технический бюллетень ВНИОРХ. - Л., 1958. - N 6-7. - С. 51-54.
12. Мишарин К.И. Естественное размножение и искусственное разведение посольского омуля на Байкале // Изв. биолого-географ. науч. иссл. ин-та при Иркутском гос. ун-те. - 1953. - Т.,14, вып. 1-4. - С. 3-132.
13. Мишарин К.И. Биологическое обоснование искусственного воспроизводства стад байкальского омуля // Теоретические основы рыбоводства.- М., 1965. - С. 168-171.
14. Никольский Г.В. Экология рыб. - М.: Высшая школа; 1974. - 367 с.
15. Подлесный А.В. Современное состояние воспроизводства омуля в оз.Байкал.//Рыб. хоз-во. - 1955. - N9. - С. 42-44.
16. Смирнов В.В., Смирова-Залуми С.С., Суханова Л.В. Микроэволюция байкальского омуля Coregonus autumnalis migratorius (Georgi). – Новосибирск: СО РАН – 2009. - С.147-174.
17. Смирнов В.В., Шумилов И.П. Омули Байкала. - Новосибирск: Наука. - 1974. - С. 112-123.
18. Соин С.Г. К вопросу о теоретических основах изучения эколого-морфологических закономерностей размножения рыб. // Экология размножения и развития рыб. М., 1980. - С. 16-28.
19. Сорокин В.Н. Условия естественного воспроизводства омуля в р.Селенге // Экология, болезни и разведение байкальского омуля. - Новосибирск, 1981. - С. 34-44.
20. Сорокин В.Н., Сорокина А.А. Воспроизводство селенгинской популяции омуля и экология ее молоди. - Тр. Лимнологического ин-та СО АН СССР, т.19. Новосибирск, 1977, с.141-155.
21. Сорокин В.Н., Сорокина А.А., Тавониус А.В. К морфологической характеристике омуля в первый год жизни. – В кн. Экологические исследования водоемов Сибири. Иркутск,1978, с.76-79.
22. Сорокин В.Н., Сорокина А.А., Михалкин А.Ф., Щербаков А.М. Характеристика нерестилищ и ската личинок северобайкальского омуля. - В кн. Озера Прибайкальского участка зоны БАМ. Новосибирск, 1981, С.185-194.
23. Шумилов И.П. Выживаемость икры байкальского омуля Coregonus. auttumnalis migratorius (G.) на нерестилищах р. Кичеры и влияние водности реки на урожайность поколений. – "Вопросы ихтиологии», 1971, т. 2, вып. 2(67), с. 280-289.
24. Щербаков А.М.Динамика ската личинок омуля с нерестилищ р.Кичеры. – Сб. Динамика продуцирования рыб Байкала., Новосибирск, Наука, 1983,.С. 141-152.
Обсуждение результатов
Функция покатной миграции личинок - обеспечение оптимальных условий их нагула (Соин, 1980), уход от потенциальных хищников, что, в конечном счете, гарантирует максимальную выживаемость. Поэтому характер и протяженность покатной миграции обеспечивает благоприятные места нагула, оптимальную плотность распределения, а это обеспеченность кормом, уход от хищника, сведение к минимуму конкуренции. В свою очередь нерестовая миграция определяет протяженность и длительность покатной миграции будущего потомства. Таким образом, потомство от производителей первого периода захода на первых этапах постэмбрионального развития оказываются в наиболее благоприятных условиях нагула, по сравнению с потомством от производителей второго и тем более третьего периодов.
Так аномально поздний заход 2016 года производителей омуля в реки Верхнюю Ангару и Кичеру и наверняка с низкими миграционными возможностями, что не позволило в полной мере реализовать требуемые дальности хода для достижения наиболее благоприятных нерестилищ.
Если рассматривать покатную миграцию личинок омуля с естественных нерестилищ Верхней Ангары за прошлый период, когда количество скатывающихся личинок находилось на уровне 1 млрд. шт. и более, то максимальные значения на кривой динамики их ската были более продолжительны во времени и зачастую имели несколько пиков. То есть интенсивность ската личинок носила как бы импульсивный характер (Шумилов, 1974; Сорокин, и др., 1981; Войтов А.А., 1981; Шербаков, 1983). Как показали наблюдения 2017 года, пусть и незначительный, но скат личинок омуля с р.Котера прошел немого раньше, т.е. до основного ската личинок с нерестилищ р.В.Ангара (19-21 мая), р.Окушанда -15 мая. Исходя из анализа данных, ранее проводимых работ следует, что в р.Верхняя Ангара вылупление и скат личинок омуля происходит при первом весеннем подъеме уровня воды, которому предшествует прогрев воздуха. Проведенные нами исследования показали, что этот комплекс факторов дифференцирован для каждого притока реки обусловленных временем наступления этих условий в распадке, где протекает этот приток. В многолетнем аспекте в бассейне реки Верхняя Ангара скат личинок происходит со второй декады мая по первую декаду июня при колебаниях на 10-15 суток. Если весна дружная и подъем уровня воды в разных притоках совпадает, в нижнем течении р.Верхняя Ангара наблюдается мощный паводок и вода выходит из берегов, что отрицательно сказывается на выживаемости скатывающихся личинках омуля. Но, как правило, паводок в разных притоках наступает поочередно от устья к истоку, т.е. с Юга на Север. Поэтому и основной скат личинок омуля в каждом притоке не совпадает, что и отражается на кривой динамики интенсивности ската.
Анализ имеющих данных и наши наблюдения показывают, что численность и динамика скатывающихся личинок омуля соответствуют численности нерестового стада производителей и особенностям их при освоении нерестилищ.
С конца 90х годов в последние два десятилетия численность скатывающихся личинок омуля в Верхняя Ангара находится на уровне очень низкой и весьма стабильной величине порядка 10 % от потенциального фонда икры заходящих производителей. Столь стабильно низкая эффективность естественного воспроизводства при разном уровне водности реки кажется сомнительной. Для байкальского омуля такая эффективность воспроизводства находится не ниже критических величин (Воронов, 2003), и она не может обеспечить пополнения, гарантирующего его воспроизводительную способность. Установлено, что для каждого вида существует зависимость между индивидуальной абсолютной плодовитостью и выживаемостью потомства. Если рассматривать фазу покатной миграции личинок в воспроизводственном цикле байкальского омуля, то для обеспечения промыслового изъятия в современных условиях на уровне 20-25 тыс. ц, то среднемноголетняя величина ската личинок с р.Селенга должна быть на уровне 1.5 млрд. шт. (Воронов, 1993), для р.Верхняя Ангара 2.2-2.8 млрд. шт. (Шумилов, 1974). Хотя для байкальского омуля и установлена зависимость (Калягин, 1996; Воронов, 1993), как и для большинства видов рыб (Никольский, 1974) - выживаемости поколения от его исходной численности. Для байкальского омуля это количества скатывающихся личинок и чем ниже ее величина, тем выше выживаемость от данного поколения, но эти величины предельны. Для омуля пелагической морфогруппы минимальная критическая величина скатывающихся личинок находятся на уровне порядка 60-70% от оптимальной и не более 3-4 лет подряд, когда сохраняется отработанный режим промысла и оптимальный уровень воспроизводства (Воронов, 1993). Ярким примером этому может служить факт срыва воспроизводства омуля на р.Селенге в 1981-89 гг., когда эффективность реализации репродукционного потенциала была снижена в 2.2 раза от оптимальной величины из-за применения электро-рыбо-заградительного устройства. В натуральном выражении средняя величина скатывающихся личинок вместе с личинками выпускаемыми с Селенгинского рыбоводного завода упала в среднем до 0.72 млрд. шт, которые обеспечили заход в среднем 1.12 млн. производителей, а от этих производителей средний скат составил 0.64 млрд. личинок, из которых средний заход в р.Селенгу составил 1.02 млн. экз. Это отрицательно сказалось и на величине промыслового изъятия пелагической морфогруппы особенно на Селенгинском мелководье – его доля в уловах омуля упала с 70-80% (в 80х годах) до 40-55% (в 90х годах) обшей добычи омуля в этом промрайоне. В этот период по данным официальной статистике произошло и снижение общего вылова омуля по Селенгинскому промрайону с 800 до 300 тонн.
Поэтому резкое снижение численности скатившихся личинок омуля с р.Верхняя Ангара в 2017 году 135 млн.шт., что почти в 10 раз ниже оптимальной величины, необходимо рассматривать как катастрофическое состояние уровня воспроизводства, которое при всех суммарных компенсаторных факторах (Калягин, 1996) не в состоянии обеспечить нормальное функционирование популяции и тем более промысел.
Анализ всех природных факторов в период ската личинок омуля в р.Верхняя Ангара в 2017 году позволяет характеризовать их, как оптимальные. Вылупление и скат личинок прошли в средние сроки для этой реки при подъеме уровня воды не превышающий среднемноголетний. Поэтому выхода воды из берегов не произошло, а, следовательно, и потери скатывающихся личинок на разливах. Оптимальные сроки выхода личинок на места нагула при небольшой их численности обеспечили темп их роста несколько выше средних значений, а это позволяет надеяться на более высокую величину выживаемости омуля от поколения этого года.
По данным Войтова А.А. заход производителей омуля реки Верхняя Ангара до 2011 года был на уровне 2.3 млн.экз., потенциальный фонд икры 8.7 млрд. икринок, скат личинок около 1 млрд.шт., с 2012 г. такой же численности заходящих производителей наблюдается резкое снижение количества скатывающихся личинок почти в 2.5 раза от потенциального фонда икры и в 1,7 раза в количественном выражении. Резкое снижение количества скатывающихся личинок омуля с 2012 года произошло и на р.Селенге (Базов, 2016), хотя до последних двух лет 2016 и 2017 годах официальные данные по количеству заходящих производителей, при некотором снижении, оставались в пределах ошибки учета. Для выяснения причин столь значительного несоответствия численности заходящих производителей – количеству скатывающихся личинок и величины промыслового изъятия требуют специального анализа.
При изучении покатной миграции личинок омуля в р.В.Ангара (Шумилов, 1974) была установлена зависимость размерно-весовых показателей личинок в течение их покатной миграции – вначале скатываются самые мелкие, затем крупнее и в конце покатной миграции самые крупные, в то время как в р.Кичера в период всего ската различий в скатывающихся личинках нет. По мнению автора увеличение размера личинок происходит вследствие их развития за период ската с верхних нерестилищ, а отсутствие такового у личинок в р.Кичеры из-за близости пункта учета от нерестилищ. При изучении покатной миграции личинок омуля в р.Селенге установлено, на постоянном створе 35 км от устья вначале покатной миграции скатываются наиболее крупные личинки затем закономерно уменьшаясь как в размерном, так и весовом составе (Афанасьев 1980). В то время как, при исследовании этих показателей на удаленных друг от друга участках реки (155 км, 35 км, 2 км от устья) при относительном сохранении длины, вес личинок достоверно уменьшался (Афанасьев, 1980; Сорокин и др., 1981). Уменьшение весовых показателей по мере ската личинок авторы объясняют затратой запаса энергии при эндогенном питании.
Размеры икринок на стадии овуляции у омуля разных морфогрупп достоверно отличаются. Так наиболее крупная икра у омуля прибрежной морфогруппы, а наиболее мелкая у омуля придонно-глубоководной, среднее положение занимает икра у омуля пелагической морфогруппы. Из более крупных икринок вылупляются и более крупные предличинки. Расовая структура нерестового стада омуля каждой реки не однородна и представлена как минимум двумя морфогруппами. Исследованиями автора, проводимые по изучению нерестовых миграций омуля в реках Байкала в частности установлено, что на р.Селенге и Верхней Ангаре вначале нерестовой миграции идут наиболее крупные и сильные производители омуля пелагической морфогруппы. Это либо повторно нерестующие, либо наиболее быстрорастущие особи.
Наши исследования покатной миграции личинок также подтверждают ранее выявленные закономерности по изменению размерных показателей личинок омуля, как в реке Селенга, так и в р.Верхняя Ангара. Только вот причина этих закономерностей, что в р.Верхняя Ангара вначале скатываются более мелкие личинки, а затем более крупные, по нашему данным, обусловлены скатом личинок омуля пелагической морфогруппы, так как они находятся на более удаленных верхних нерестилищах, а затем личинки прибрежной морфогруппы. Вскрытие реки Селенга и первый весенний паводок в р Верхняя Ангара начинается с верховья каждого притока, поэтому вылупление и скат личинок начинается с наиболее верхних участков нерестилищ. В р.Верхняя Ангара вначале покатной миграции скатываются личинки омуля пелагической морфогруппы, которые более мелкие, а затем личинки омуля прибрежной морфогруппы, которые крупнее пелагических. А в р.Селенга вначале скатываются более крупные личинки от элитной части производителей пелагической морфргуппы, а затем с ниже расположенных нерестилищ от остальной части производителей, т.е. по мере вскрытия реки, а также ската личинок от придонно-глубоководной морфогруппы, которые мельче пелагических.
Из вышесказанного следует, что покатная миграция личинок омуля в каждой реке это экологически обусловленный процесс на этапе воспроизводства, как адаптивный фактор, направленный на обеспечение выживаемости с учетом особенностей воспроизводства в конкретных условиях каждой реки.
То, что сроки выклупления и характер ската личинок омуля экологически выдержаны и обусловлены, объясняет задержку скатившихся личинок омуля с нерестилищ Верхней Ангары в водоемах верхней части Северобайкальского сора. До прихода второй основной волны весеннего паводка большая Северо-Восточная часть сора была слишком мелководной (менее 50 см) с выраженным речным режимом основных проток выхода в открытый Байкал, который к тому же еще был покрыт льдом. В этот период условия для нагула личинок омуля неблагоприятны. С приходом второй волны паводка с 10 по 14 июня 2017 года условия в этой части сора стали благоприятными – поднялся уровень воды, скорость течения выноса воды из сора в озеро по основным прорвам снизилась, Байкал освободился ото льда. Непосредственно около косы и вдоль Восточного берега Байкала тоже образовалась зона теплой речной воды.
Для установления более четких и обусловленных закономерностей динамики выхода личинок из прибрежно-соровой системы в открытую часть Северного Байкала, проследить их дальнейшую миграцию и условия нагула требуются специальные исследования. Наблюдения, проведенные на фиксированных разрезах в прибрежной зоне Северной оконечности Байкала (Шумилов, 1974; Сорокин, 1977) свидетельствуют об увеличении разброса линейно-весовых показателей нагульных личинок омуля от июня к июлю. Эти факты указывают на достаточно сложный механизм приспособлений личинок омуля к разным условиям нагула в этой экосистеме. С одной стороны есть основания предполагать и наши исследования это подтверждают, что вылупление личинок на нижних участках нерестилищ происходит и в июне уже после основного ската личинок. Кроме этого личинки, а затем и молодь омуля кичерской популяции далеких миграций не совершает и продолжает нагул на северном Байкале (Смирнов, 2009), в то время как личинки и молодь ангарской популяции на нагул уходит на Юг Байкала, совершая миграции в прибрежной зоне вокруг озера. Это подтверждается тем, что уже весной в Малом Море нагуливаются годовики омуля пелагической и прибрежной морфогрупп.
Выводы
1. Основной скат личинок омуля с нерестилищ бассейна реки Верхняя Ангара в 2017 году прошел в период первого весеннего паводка с 18 по 21 мая при подъеме уровня на 194см, падении прозрачности с 3,5 м до 0.25 м, понижении температуры воды с 10.3ºС до 3.5-6ºС эти условия следует считать периодом экологического оптимума.
2. Аномально поздний заход производителей омуля 2016 года при костной нормативно-правовой базе способствовали официальному вылову производителей идущих на нерест, в результате реализованный репродукционный потенциал зашедших производителей составил 25.3%.
3. При позднем заходе производителей омуля в р.Верхняя Ангара, р.Окушанда играет существенную роль в воспроизводстве омуля, однако условия инкубации икры в 2016-17 году были нарушены и основная часть икры погибла, скат личинок составил 2.65 млн. личинок, а снос учтенных мертвых икринок150.89 млн. шт.
4. Личинки омуля из р.Кичера сразу скатываются в Северо-Западную часть Северобайкальского сора, а затем с потоком теплой соровой воды распространяется вдоль СЗ побережья оз.Байкал, освобождающуюся ото льда. Покатные личинки омуля с р.Верхней Ангары задерживаются в верховьях СВ части сора и позже при втором более мощном весеннем паводке (в 2017 году 10-14 июня), уже в более развитом состоянии выходят в СВ часть сора и с потоком теплых соровых вод выходят в прибрежную часть Северного Байкала, к этому времени освободившуюся ото льда.
5. Сроки вылупления и покатную миграцию личинок омуля экологически выдержанных и обусловленных следует рассматривать как один из элементов функциональных адаптаций в воспроизводственном цикле байкальского омуля.
6. Для установления более четких закономерностей динамики выхода и нагула личинок из прибрежно-соровой системы в открытой части Северного Байкала и дальнейших их миграций требуются специальные исследования.
Список литературы:
1. Афанасьев Г.А. Экология и воспроизводство байкальского омуля в реке Селенге: Автореф. дис. канд. биол. наук. - Иркутск, 1980. - 23 с.
2. Афанасьев Г.А. Экология нерестового стада омуля р.Селенги // Экология, болезни и разведение байкальского омуля. - Новосибирск, 1981.- С. 5-34.
3. Базов А.В., Базова Н.В. Селенгинская популяция байкальского омуля: прошлое, настоящее, будущее. – Улан-Удэ6 Изд-во БНЦ СО РАН, 2016, 352 с.
4. Войтов А.А. Биологическая характеристика и воспроизводство омуля в р.Верхняя Ангара. // Экология, болезни и разведение байкальского омуля. - Новосибирск, 1981.- С. 75-82.
5. Воронов М.Г. Эколого-биологические основы повышения эффективности воспроизводства омуля в р.Селенге в современных условиях.// Автореферат диссертации на соискание ученой степени кандидата биологических наук. - Санкт-Петербург, 1993. - 18с.
6. Воронов М.Г. Разнообразие байкальского омуля, структурная организация воспроизводящей части популяции и проблемы их сохранения.// Первый конгресс ихтиологов России: Тез. Докл. сентябрь г.Астрахань. – М., ВНИРО, 1997, С.
7. Воронов М.Г. К вопросу о перспективах искусственного воспроизводства байкальского омуля. Тез.докл. г.Улан-Удэ, 2003, С
8. Гербильский Н.Л. Биологическое значение и функциональная детерминация миграционного поведения анадромных рыб // Биологическое значение и функциональный детерминизм миграционного поведения животных. - М.; Л., 1965. - С. 18-33.
9. Дормидонтов А.С. Нерест и вопросы ленской ряпушки // Вопросы рыбного хозяйства Восточной Сибири. - Иркутск, 1969. - С. 9-19.
10. Дормидонтов А.С. Нерестовые миграции как адаптации, обеспечивающие расселение сигов по местам нагула // Зоологические исследования Сибири и Дальнего Востока: V Всесоюз. симпоз. Биологич.проблемы Севера, 1972, Магадан. - Владивосток, 1974. - С. 164 -168.
11. Краснощеков С.И. О биологии личинок байкальского омуля. //Тр./ Научно-технический бюллетень ВНИОРХ. - Л., 1958. - N 6-7. - С. 51-54.
12. Мишарин К.И. Естественное размножение и искусственное разведение посольского омуля на Байкале // Изв. биолого-географ. науч. иссл. ин-та при Иркутском гос. ун-те. - 1953. - Т.,14, вып. 1-4. - С. 3-132.
13. Мишарин К.И. Биологическое обоснование искусственного воспроизводства стад байкальского омуля // Теоретические основы рыбоводства.- М., 1965. - С. 168-171.
14. Никольский Г.В. Экология рыб. - М.: Высшая школа; 1974. - 367 с.
15. Подлесный А.В. Современное состояние воспроизводства омуля в оз.Байкал.//Рыб. хоз-во. - 1955. - N9. - С. 42-44.
16. Смирнов В.В., Смирова-Залуми С.С., Суханова Л.В. Микроэволюция байкальского омуля Coregonus autumnalis migratorius (Georgi). – Новосибирск: СО РАН – 2009. - С.147-174.
17. Смирнов В.В., Шумилов И.П. Омули Байкала. - Новосибирск: Наука. - 1974. - С. 112-123.
18. Соин С.Г. К вопросу о теоретических основах изучения эколого-морфологических закономерностей размножения рыб. // Экология размножения и развития рыб. М., 1980. - С. 16-28.
19. Сорокин В.Н. Условия естественного воспроизводства омуля в р.Селенге // Экология, болезни и разведение байкальского омуля. - Новосибирск, 1981. - С. 34-44.
20. Сорокин В.Н., Сорокина А.А. Воспроизводство селенгинской популяции омуля и экология ее молоди. - Тр. Лимнологического ин-та СО АН СССР, т.19. Новосибирск, 1977, с.141-155.
21. Сорокин В.Н., Сорокина А.А., Тавониус А.В. К морфологической характеристике омуля в первый год жизни. – В кн. Экологические исследования водоемов Сибири. Иркутск,1978, с.76-79.
22. Сорокин В.Н., Сорокина А.А., Михалкин А.Ф., Щербаков А.М. Характеристика нерестилищ и ската личинок северобайкальского омуля. - В кн. Озера Прибайкальского участка зоны БАМ. Новосибирск, 1981, С.185-194.
23. Шумилов И.П. Выживаемость икры байкальского омуля Coregonus. auttumnalis migratorius (G.) на нерестилищах р. Кичеры и влияние водности реки на урожайность поколений. – "Вопросы ихтиологии», 1971, т. 2, вып. 2(67), с. 280-289.
24. Щербаков А.М.Динамика ската личинок омуля с нерестилищ р.Кичеры. – Сб. Динамика продуцирования рыб Байкала., Новосибирск, Наука, 1983,.С. 141-152.

Свой бизнес
по разведению ценных пород рыбы в УЗВ
Нажимая на кнопку, вы даете согласие на обработку персональных данных и соглашаетесь c политикой конфиденциальности